Working Papers in Neurolinguistics and Neuroscience |
|
Neuro 1: |
Michael Schecker: Neuronale "Kodierung" zentraler Sprachverarbeitungsprozesse (1998) |
(anläßlich Friedemann Pulvermüller, "Neurobiologie der
Sprache",
Pabst Science Publishers, Psychologia Universalis - Neue Reihe: Bd. 1, 1996)
-----------
(1) Einleitung
-----------
Die folgenden Ausführungen sind angeregt durch Pulvermüllers Habilitationsschrift "Neurobiologie der Sprache". Was ist das für ein Buch? Vorweg: Es ist ein wichtiges Buch, es ist ein faszinierendes Buch, es ist ein ärgerliches Buch:
1.1. Pulvermüller etabliert und diskutiert die Vorstellung, daß Sprache, daß sprachliche Segmente, daß Prozesse der Sprachverarbeitung im wesentlichen auf Zell-Verbänden beruhen (Theorie der "Cell Assemblies"), deren interne Vernetzung über Lernprozesse erworben wird;
1.2. er wendet die Theorie der "Cell Assemblies" - u.a. unter Rückgriff auf ein künstliches neuronales Netz (ohne jedwede funktionale Architektur) - auf aphasische Syndrome unterschiedlicher Art an;
und diskutiert ausführlich -
1.3.1. anhand lateralisierter tachistoskopischer Präsentationen (Wort-Entscheidungsaufgaben, Erhebung der Reaktionszeiten) und
1.3.2. anhand der Erhebung Ereignis-korrelierter Potentiale (EKP-Studien, wobei er u.a. die Lokalisierungsproblematik einfach unterschlägt), hier auch der Auswertung der sogenannten 'Gamma- Band-Aktivität';
1.3.3. anhand von Magnetoenzephalogramm-Auswertungen (MEG)
die unterschiedliche Verarbeitung von sogenannten Funktionswörtern und sogenannten Inhaltswörtern (die - entgegen den Ausführungen Pulvermüllers - linguistisch natürlich keine Wortarten darstellen);
1.4. und entwickelt schließlich und endlich weitestgehende Ausblicke und Erklärungsansätze für fast alles, was in der Sprachwissenschaft und in den Neurowissenschaften derzeit an interessanten und relevanten Fragestellungen bearbeitet wird. (Die Schlichtheit der Überschrift des abschließenden Kapitels überzeugt: "Sprachwissenschaftliche Fragen und ihre gehirntheoretischen Antworten"!!!)
Und das alles auf rund 116 minus 11 Seiten Text (die Darstellung selber beginnt erst auf Seite 12)!
-----------
2.1. Pulvermüller führt - mit Recht - gegen eine Vielzahl von Ansätzen der modernen Linguistik und der Neurowissenschaften aus, daß hier mit Modellvorstellungen gearbeitet werde, "die nicht explizit auf Gehirnstrukturen Bezug nehmen"; eine der Kapitelüberschriften Pulvermüllers lautet: "Module statt Neuronen".
2.2. Der entscheidende Punkt ist aber nicht die Frage, ob die Sprachverarbeitung modular aufgebaut ist (dazu auch weiter unten), sondern ob für die Modellierung der Sprachverarbeitung netzwerktheoretische oder an der Arbeitsweise von Computern orientierte Modelle zugrundegelegt werden. Hier gleichen sich so unterschiedliche Modelle wie das Sprachproduktionsmodell von Garrett und dasjenige von Levelt: Beide Modelle sind an der Arbeitsweise von Computern orientiert; und das Gehirn arbeitet ganz und gar nicht wie ein Computer!
Was kennzeichnet solche an der Arbeitsweise von Computern orientierten Modellvorstellungen? Warum handelt es sich hier um Modelle der menschlichen Sprachverarbeitung, zu denen das Original gewissermaßen fehlt (eben weil das menschliche Gehirn grundsätzlich anders arbeitet)?
2.3. Bei Garrett und Levelt und vielen anderen werden Informationen in Speichern (z.B. im Lexikon) gespeichert - und je nach Anforderungen (von Prozessoren?) aufgerufen und weiterverarbeitet. Weiterverarbeitung bedeutet darüberhinaus, daß die Information sich qualitativ verändert; an modularen Modellen orientiert unterscheiden sich pro Modul oder Verarbeitungskomponente input und output eines Moduls bzw. einer jeweiligen Verarbeitungskomponente.
Dazu ein für sich sprechendes Detail: Damit der Aufruf einer gespeicherten Information gelingen kann, muß diese Information auf einem 'adressierten Speicherplatz' zur Verfügung stehen; wie sonst wüßte ein Prozessor, wo er die gerade benötigte Information finden kann? Ganz entsprechend sieht Levelt im Rahmen seines Inhalts- oder Konzepte-Lexikons vor, daß bereits dort auch eine Adresse für eine erst später benötigte Wortform aus dem Wortformen-Lexikon gespeichert ist.
2.4. Im Gehirn gibt es nun freilich keine Speicher mit adressierten Plätzen (es sei denn, man will das Ganze - höchst vage, höchst willkürlich - als irgendeine Art von Metaphorik - Metaphorik für was? - verstehen); allgemeiner läßt sich im Gehirn zwischen Speichern und Prozessoren nicht unterscheiden. Und schließlich finden zumindest in den zentralen Verarbeitungsprozessen des Gehirns (dazu gleich noch mehr) auch keine Datentransporte statt; sondern hier haben wir es der Qualität nach mit stets gleichen exzitatorischen wie inhibitorischen Aktivationen auf einem neuronalen Netzwerk zu tun.
2.5. "Der modularistische Ansatz läßt die Antwort auf all diese Fragen offen", schreibt Pulvermüller - und nicht mehr! Das bedeutet eine völlig unzulässige Verharmlosung: Modelle, die an der Arbeitsweise von Computern orientiert sind und u.a. mit Speichern und Prozessoren arbeiten, können prinzipiell keine Modelle der Arbeitsweise des Gehirns sein!
Im übrigen trifft die Kritik nicht unbedingt modularistische Ansätze: Auch im Rahmen von Netzwerkmodellen wird von Fall zu Fall modularistisch konzipiert.
-----------
3.1. In den Überlegungen der modernen Neurobiologie hat die Einsicht Einzug gehalten, daß wir für das Gehirn zwischen periphären und zentralen Prozessen der Informationsverarbeitung unterscheiden müssen: Nervenzellen brauchen - nachdem sie einmal aktiviert waren - eine gewisse Zeit (absolute und relative Refraktärphase), bis sie wieder aktiviert werden können; rechnet man die Zeit hinzu, die es braucht, bis eine überschwellige Aktivation entstanden ist, so macht das deutlich, daß eine Nervenzelle nur mit einer bestimmten maximalen Frequenz feuern kann; real beobachtbar sind im Rahmen periphärer Verarbeitungsprozesse bis zu 100 HZ, im Rahmen zentraler Informationsverarbeitungsprozesse aber nur um etwa 10 HZ.
3.2. Eine Vielzahl von neurobiologischen Studien hat belegt, daß in periphären Prozessen frequenzkodiert wird: Ein gutes Stück Informationsgehalt steckt hier in der Zahl der Aktivationen oder 'Spikes', die als 'burst', als Bündel aufeinander folgender Impulse, von neuronaler Position zu neuronaler Position weitergegeben werden. Gut belegt ist Frequenzkodierung für primäre visuell-sensorische, aber auch für primär auditiv-sensorische Informationsverarbeitung.
3.3. Legen wir für zentrale Verarbeitungsprozesse (wo immer im Detail die Grenze liegt) Frequenzen von rund 10 HZ zugrunde und bedenken wir weiter, mit welcher Geschwindigkeit ein native speaker Sprache auf einer Vielzahl von Verarbeitungsebenen verarbeitet, so können zentrale Sprachverarbeitungsprozesse nicht frequenzkodiert sein: Bei durchschnittlich 10 Hz würde die Verarbeitung frequenzkodierter Information, würden 'Aktivationsburst's' derart viel Zeit verbrauchen, daß die für natürliche Sprachen typischen Geschwindigkeiten nicht einmal näherungsweise erreicht werden könnten.
Bei 'burst's' bzw. Impulsraten z.B. von 40 Spikes würde das im Rahmen zentraler Verarbeitungsprozesse pro 'burst' 4 sec. ergeben! Und das ist viel zu langsam: Was etwa die Phonemebene angeht (das ist immer noch der Bereich zentraler Informationsverarbeitungsprozesse), so können wir - wenn wir schnell sprechen - pro Phonem bei einfachen Konsonanten und bei Kurzvokalen ohne Probleme auf etwa 65 msec heruntergehen bzw. rund 15 Phoneme pro Sekunde artikulieren und rezipieren.
3.4. Die für 'realistische' Modelle der Sprachverbeitung zentrale Unterscheidung von periphären und zentralen Prozessen fehlt bei Pulvermüller (und das ist nicht das einzige, was an zentralen Details fehlt - vgl. dazu auch noch weiter unten; entsprechend reduziert, ja 'ausgemergelt', wirken Pulvermüllers Modellvorstellungen). Statt dessen wird mit Daten aus der Sehphysiologie (periphäre Verarbeitung) und der primären auditiv-sensorischen Verarbeitung (ebenfalls periphäre Prozesse) für Modellüberlegungen zur zentralen Informationsverarbeitung argumentiert!
-----------
4.1. Bleibt nach allem für zentrale Prozesse der Sprachverarbeitung nur die Annahme, daß hier EinzelImpuls-Verarbeitung vorliegt. Da die einzelnen Impulse oder Aktivationen jedoch stets gleich verlaufen, steckt dabei die Information - die ganze Information! - nicht in der jeweiligen neuronalen Aktivation bzw. im einzelnen Impuls, sondern ausschließlich in der Vernetzung der Nervenzellen bzw. in den Bahnen, die solche aufeinander folgenden Aktivationen auf dem neuronalen Netzwerk des Gehirns nehmen. Anders ausgedrückt steckt der Informationsgehalt im Bereich zentraler Informationsverarbeitungsprozesse ganz und gar in der Architektur des Netzwerks und der darauf über Lernprozesse installierten Bahnungen (ich komme darauf zurück).
4.2. Einzel-Impuls-Verarbeitung ist in verschiedenen Formen denkbar. Neben der Vorstellung, daß einzelne Neuronen - dann wohl mit einer beträchtlichen Redundanz - an nachgeschaltete Neuronen Impulse bzw. Aktivationen weitergeben ('Großmutterzellentheorie' - dazu gleich noch mehr), steht die Vorstellung 'verteilter Systeme', in denen modulartig organisierte bzw. vernetzte Neuronenpopulationen Impulse untereinander austauschen; für solche Vorstellungen verwenden Pulvermüller und die von ihm zitierten Autoren den Terminus "Cell Assembly".
Ein "Cell Assembly" - so Pulvermüller - ist "eine Gruppe cortikaler Neuronen [warum nur kortikal, warum nicht auch subcortikal? - eine Frage, die uns Pulvermüller schuldig bleibt], die untereinander stark exzitatorisch verknüpft sind, wobei sich die starke Verschaltung aufgrund häufiger gemeinsamer Aktivität ausgebildet hat [ ... ]. Die starke exzitatorische Verschaltung bedingt, daß die Mitglieder einer Assembly, die Assembly-Neuronen, sich gegenseitig stark beeinflussen. Wenn ein genügend großer Teil der Neuronen aktiv ist, so wird deshalb nach kurzer Zeit die gesamte Assembly aktiviert werden." (S. 24)
4.3. Pulvermüller argumentiert gegen die "Großmutterzellentheorie", daß ein solches System u.a. sehr störungsanfällig sei. Das ist jedoch ein Argument, das nur zutrifft, wenn hier nicht zusätzlich Redundanz angenommen wird. Unter Einbezug von Redundanz würde eine Läsion auch hier nicht notwendig zum Ausfall der so erbrachten Leistung insgesamt führen.
4.4. Es ist derzeitig nicht möglich, empirisch zwischen der 'Großmutterzellentheorie'
und "Cell Assemblies" zu entscheiden. Vielleicht ist das aber auch gar nicht von
zentralerer Bedeutung; vielleicht ist sehr viel zentraler die Einsicht, daß man - egal ob
im Rahmen einer 'Großmutterzellentheorie' oder auf der Basis von "Cell
Assemblies" - ohne eine funktionale Architektur mit spezifischen Teilleistungen und
deren interner Vernetzung nicht auskommt.
Und hier leisten nun allerdings trotz des generellen Zuschnitts Sprachproduktionsmodelle
wie das von Garrett oder Levelt erheblich mehr als die von Pulvermüller angestellten
Überlegungen.
4.5. Anders formuliert: Was ist eigentlich gewonnen, wenn man feststellt, daß "Cell Assemblies" - und nicht etwa 'Großmutterzellen' - die neuronalen Repräsentanten von Merkmalen, Phonemen, Wortformen, Bedeutungen usw. usw. usw. sind? Herzlich wenig! Nämlich, daß Inhaltswörter bei einer fokalen Läsion in aller Regel deshalb weniger stark betroffen sind, weil die sie repräsentierenden "Cell Assemblies" räumlich weiter streuen. (Das Gleiche ließe sich - bei Einbezug von Redundanz - auch im Rahmen der 'Großmutterzellentheorie' formulieren.)
Was an den zentralen Erkenntnissen Pulvermüllers vor allem beim zweiten oder gar dritten Lesen so bestürzt, ist der Umstand, daß sie sich - die zentralen Erkenntnisse Pulvermüllers - letztlich in solchen Feststellungen erschöpfen. Ein sprechendes Beispiel für den spezifischen Erkenntnisgewinn, den man als Leser mit Pulvermüller hat, ist der Einbezug zeitlicher Strukturierungen. Hier erschöpft sich Pulvermüller letztlich in der Feststellung, daß offensichtlich Neuronen nicht nur gleichzeitig, sondern auch zeitlich hintereinander aktiviert sein können: Wie wahr!
-----------
5.1. Wie könnte eine neurofunktionale Architektur der oben angemahnten Art aussehen (wobei es hier nur um eine grobe Skizze gehen kann)? Im folgenden konzentriere ich mich strikt auf Sprachverarbeitungsprozesse (und gehe, wie auch Pulvermüller, davon aus, daß Sprachverarbeitungsprozesse - wie keine kognitive Leistung sonst - Prinzipien und funktionale Architektur des Gehirns insgesamt spiegeln). Im einzelnen stützen wir uns am Neurolinguistischen Labor auf Beobachtungen zum natürlichen Spracherwerb, zu alltäglichen Sprachproduktions- und Sprachrezeptionsprozessen Erwachsener und auf Beobachtungen zu einer Reihe von Sprachpathologien.
Alle folgenden Überlegungen sind im Sinne der 'Großmutterzellentheorie' zu verstehen, lassen sich jedoch problemlos ohne Veränderungen als 'verteilte Systeme' bzw. auf der Basis von "Cell Assemblies" reformulieren.
Wer sich im Detail über die am Neurolinguistischen Labor der Universität Freiburg
entwickelten Vorstellungen einer neurobiologisch präzisierten oder doch präzisierbaren
neurofunktionalen Architektur informieren möchte, der sei auf Günter Kochendörfer:
Neuronale Modelle des Sprachverstehens, "cognito 2" ,
Freiburg: Hochschul-
Verlag 1997, und auf Günter Kochendörfer, Michael Schecker: Schizophrenie,
Psychopharmaka, Stottern, "cognitio
3" , Freiburg: HochschulVerlag 1998, verwiesen.
5.2. Am Beispiel der Rezeption gesprochener Sprache geht es
- (a) auf akustisch-auditive Merkmalsanalysen folgend (= periphäre Verarbeitungsprozesse)
- (b) 'bottom-up' um die Mustererkennung u.a. von Sprachlauten (Ergebnis könnten so etwas
wie Phoneme sein) und um deren 'Instantiierung' u.a. im Rahmen zeitlich-sequentiell
organisierter Wortformen (hier handelt es sich bereits um zentrale
Sprachverarbeitungsprozesse).
(c) Wortformen stellen die erste Stufe einer ganzen Hierarchie sequenzieller Verarbeitungsprozesse dar (wobei 'Sequenz' hier immer 'zeitliche Sequenz' meint).
(d) Der Input (das betrifft die Hierarchie-niedrigste Sequenz) bzw. bottom-up-Informationen können lückenhaft sein; es kann sich - in einem sehr weiten Sinne des Wortes - um uneindeutige Informationen handeln; der Input kann - bezogen auf die zur Verarbeitung anstehende Sequenz - fehlerhaft sein: Für solche und andere Fälle gehen wir über die bisher skizzierte 'bottom-up-Verarbeitung' hinaus von 'top-down-Kontrollprozessen' aus, die zumindest ein gutes Stück weit Inkohärenzen feststellen und Korrekturen auslösen (wir sprechen hier auch von prädiktiven Systemen, - im folgenden einbezogen wird allerdings nur das 'unspezifische prädiktive System').
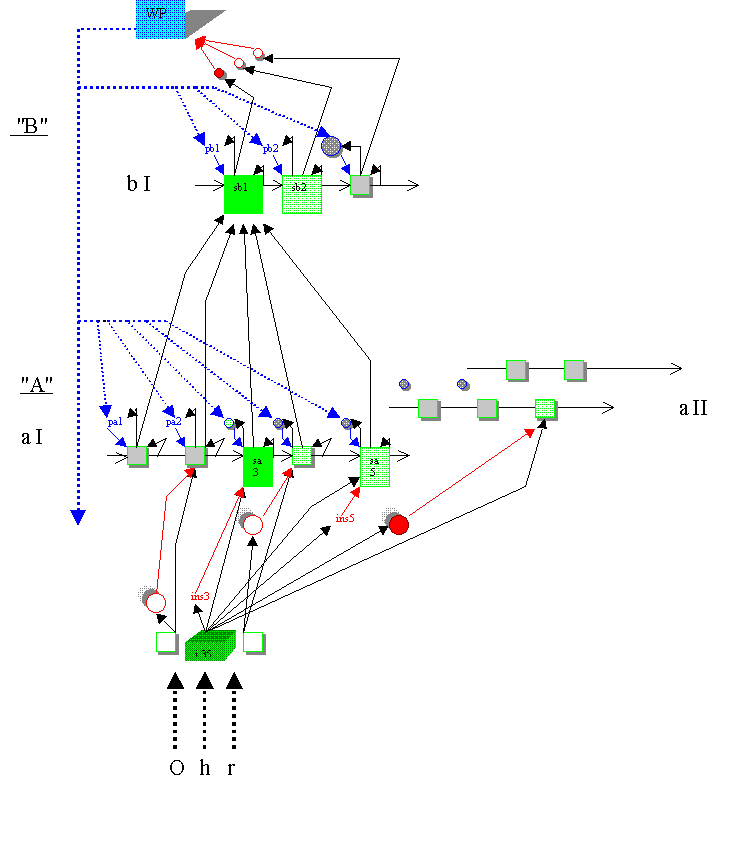
5.3. Wie funktioniert eine solche Architektur? Man vergleiche dazu die folgende Graphik:
5.3.1. Geht man für ein Zeitintervall
An einem Beispiel - sei die Sequenz 'aI' die als sequenzielles Muster gespeicherte
Phonemfolge / p a p a /; hier könnte vom ersten / p / auf das erste / a / ein Impuls
abgegeben worden sein; wird nun innerhalb eines Zeitfensters
5.3.2. Nun erzeugt ein Input 'i35' gegebenenfalls nicht nur ein unterschwelliges
exzitatorisches Potential auf 'sa3', sondern gegebenenfalls auch auf einer weiteren
sequentiellen Position 'sa5' der gleichen Sequenz 'aI' (das wäre eine zweite 'Instanz'
derselben Input-Information 'i35'). Und es ist durchaus auch denkbar, daß weitere
Instanzen von 'i35' - z.B. in einer Sequenz 'aII' - angesprochen werden. - Am Beispiel von
'/ p a p a /' werden wir den Input '/ a /' nicht nur auf das erste sequentielle '/ a /',
sondern auch auf das zweite '/ a /' beziehen. Dennoch kommt es innerhalb des
Zeitfensters
5.3.3. Ein (unterschwelliges) EPSP braucht Zeit, bis es auf natürliche Weise wieder abgebaut ist. Und zwar zuviel Zeit: Bei normal schnellem Sprechen und Hörverstehen haben wir 'sa5' gegebenenfalls bereits zu einem Zeitpunkt erreicht, zu dem noch nennenswerte Reste eines zuvor aufgebauten (unterschwelligen) EPSP's dort anliegen. Und das würde dann einmal aufgrund des eintreffenden sequentiellen Impulses von 'sa4' auf 'sa5' und zum anderen aufgrund eines dort noch anliegenden vorgängigen EPSP's zu einer vollen Aktivation führen, ohne daß notwendigerweise ein zu 'sa5' passender Input vorliegt.
Um solche und ähnliche 'assoziativen Entgleisungen' bzw. fehl laufenden Aktivationen zu vermeiden, ist es notwendig, einen Hemmmechanismus einzuführen: Derselbe Input-Impuls 'i35', der u.a. 'sa5' unterschwellig aktiviert, aktiviert auch den Hemmmechanismus 'ins5', eine Zelle oder eine neuronale Population, die um nur wenige Millisekunden zeitlich versetzt die sequentielle Position 'sa5' (wenn 'sa3' bis dahin nicht schon voll aktiviert wurde, auch 'sa3') inhibiert und das dort noch anstehende EPSP wieder auf den Ausgangswert zurückführt.
Vgl. zur Rolle, die eine Störung dieses Hemmmechanismus bei schizophrenen Psychosen spielen könnte, die Ausführungen in Günter Kochendörfer, Michael Schecker, Assoziationen und Kontextverarbeitung. Ansätze zu einer Simulation schizophrenen Sprachverhaltens. In: Gert Rickheit (Hg.), Studien zur klinischen Linguistik, Opladen (Westdeutscher Verlag) 1997, 49 - 65.
5.3.4. Sequentielle Positionen sind auf sich selber rückgekoppelt, - warum? Erneut am Beispiel einer Phonemfolge: Wenn im Rahmen der musterhaft gespeicherten Phonemfolge / S l a: f / (= "Schlaf") das lange / a: / mit Input versorgt wird, dann handelt es sich dabei zunächst einmal um ein über eine gewisse Zeitspanne hinweg anstehendes kontinuierliches Schallereignis, das jedoch - so nehmen wir an - im Rahmen der Mustererkennung zu diskret aufeinander folgenden Aktivationen ein- und derselben Input-Kategorie führt. Handelt es sich dabei z.B. um 'i35', dann führt das gegebenenfalls zur Aktivation der sequentiellen Position 'sa3', die auf sich selber rückgekoppelt (ein in seiner Auswirkung unterschwelliger Impuls) solange immer wieder voll aktiviert wird, wie 'i3' in hinreichend kurzen Abständen feuert; eben das ist unser Vorschlag zur neurofunktionalen Modellierung der Verarbeitung u.a. von Langvokalen.
Der oben beschriebene Mechanismus ist von grundlegender Bedeutung: Eine übergeordnete sequentielle Position 'sb1', die - weil Sequenzposition - auf sich selber rückgekoppelt ist, wird auch ohne einen sequentiellen Impuls vom Sequenzvorgänger aus solange immer wieder feuern, wie die mit 'sb1' vernetzten untergeordneten Sequenzpositionen 'sa1', 'sa2', 'sa3' usw. feuern. Eben das ist unser Vorschlag zu einer neurofunktionalen Modellierung der Verarbeitung z.B. syntaktischer Konstituenten.
5.3.5. Wenn eine sequentielle Position - z.B. 'sa3' - überschwellig aktiviert wird und feuert, dann - so haben unsere Ausführungen oben bisher ergeben - dann gibt sie, dann gibt 'sa3' einmal einen (in seiner Wirkung unterschwelligen) Impuls an den Sequenznachfolger, an 'sa4', ab; zum anderen ist 'sa3' auf sich selber rückgekoppelt und gibt entsprechend auch an sich selber einen (in seiner Wirkung ebenfalls unterschwelligen) Impuls ab. Und da die sequentiellen Positionen einer untergeordneten Sequenz 'aI' allesamt auch mit einer Position - z.B. mit 'sb1' - einer übergeordneten Sequenz 'bI' vernetzt sind, wird immer auch ein (in seiner Wirkung erneut nur unterschwelliger) Impuls an eine zugehörige übergeordnete Sequenzposition, an 'sb1', abgegeben.
Fehlt dieser Impuls an eine übergeordnete Sequenzposition, gibt eine sequentielle
Position 'sa3' keinen Impuls an 'sb1' ab (z.B. weil bei 'sa3' innerhalb eines
Zeitfenster
Was kann das für Auswirkungen haben? Nehmen wir an, 'sb2' haben keinen inhibierenden Impuls anf 'wp' gesendet und habe damit die 'wp-Aktivität' freigesetzt. Dann wird top-down u.a. auf die 'prädiktiven' Positionen 'pb1' und 'pb2' ein (in seiner Wirkung unterschwelliger) Inpuls abgegeben. Gehen wir nun aber davon aus, daß die sequentielle Position 'sb1' gerade gefeuert hatte - und dabei auch einen (in seiner Wirkung ebenfalls unterschwelligen) Impuls an 'pb1' abgegeben hat, dann werden diese beiden Impulse jetzt aufsummert, und 'pb1' wird voll aktiviert und feuert u.a. auf 'sb1', das auf diese Weise potentiell neu gestartet wird.
5.3.6. 'Neustarts' der hier skizzierten Art führen gegebenenfalls dazu, daß eine Sequenz 'bI' trotz überdehnter Pausen korrekt weitergeführt werden kann. Und sie führen noch zu weiteren Effekten: Ein Neustart von 'sb1' kann auch während der Abarbeitung der Sequenz 'aI' stattfinden. Wenn hier nämlich die Verarbeitung zum Erliegen kommt, dann beginnt 'sb1' gewissermažen zu schweigen - und löst auf diese Weise die 'wp-Aktivität' aus, die - diesmal in Kombination mit der Rückkopplung von 'sb1' auf sich selber - zu einem Neustart von 'sb1' führt.
Ein Fall wie der zuletzt beschriebene kann auftreten, wenn z.B. der auf die Sequenz
'aI' zu beziehende Input für die sequentiellen Positionen 'sa2' oder/und 'sa3' oder/und
'sa4' oder/und 'sa5' so 'ausgedünnt' ist, daß hier die sequentielle Verarbeitung zum
Erliegen kommt. Wenn jetzt aber 'sb1' erneut gestartet wird - und innerhalb eines
Zeitfensters
5.4. Wir haben die oben skizzierte neurofunktionale Architektur in ein künstliches Netzwerk übersetzt und damit eine Reihe natürlicher Sprachlern- und Sprachverarbeitungsprozesse erfolgreich simulieren können. Sicher sehr viel spektakulärer dürfte es jedoch sein, daß es uns auch gelungen ist, in Ausschnitten Pathologien auf dem PC zu simulieren (so 'assoziative Entgleisungen' bei Schizophrenie und spezifische Gedächtnisprobleme, wie sie bereits relativ früh bei einer Alzheimerschen Erkrankung auftreten); in diesem Zusammenhang dürften auch die Simulationen der Wirkung von Medikamenten wie Haloperidol und Clozapin bei Schizophrenie oder von Tacrin bei Alzheimerscher Erkrankung auf breiteres Interesse stoßen.
-----------
Zum Abschluß noch ein Blick zurück auf die Habilitationsschrift Pulvermüllers: Es ist ein für die Diskussion wichtiges Buch! Und wenn man nur den neurowissenschaftlichen Kenntnisstand der traditionellen Aphasiologie und die dort immer noch im Zentrum stehenden 'black-box-Modelle' in Rechnung stellt, dann ist es sicherlich auch ein faszinierendes Buch.
-----------
M. Garrett, The organization of processing structures for language production. In: D. Caplan, A.R. Lecours, A. Smith (Hgs.), Biological perspectives on language. Cambridge/Ma (MIT Press) 1984, 172 - 193
G. Kochendörfer: Neuronale Modelle des Sprachverstehens, "cognitio 2" , Freiburg: HochschulVerlag 1997
G. Kochendörfer, M. Schecker, Assoziationen und Kontextverarbeitung. Ansätze zu einer Simulation schizophrenen Sprachverhaltens. In: Gert Rickheit (Hg.), Studien zur klinischen Linguistik, Opladen (Westdeutscher Verlag) 1997, 49 - 65
G. Kochendörfer, M. Schecker: Schizophrenie, Psychopharmaka, Stottern, "cognitio 3" ,
Freiburg:
HochschulVerlag 1998
W.J.M. Levelt, Speaking: From intention to articulation. Cambridge/Ma (MIT Press) 1989
M. Schecker, Neurolinguistische Vorstellungen zur Sprachverarbeitung. Probleme und Perspektiven. In: ARBA 1, Basel 1993, 251 - 264
Michael Schecker
-----------
(Copyright "Neurolinguistisches Labor", Freiburg im Februar 1989)